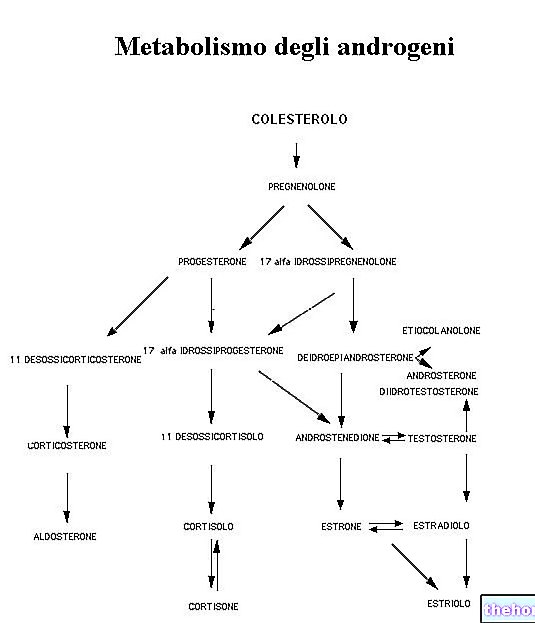
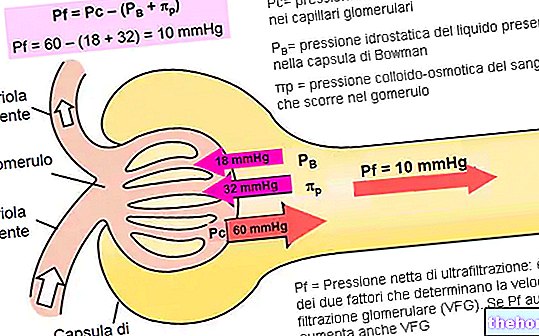
"prvi del
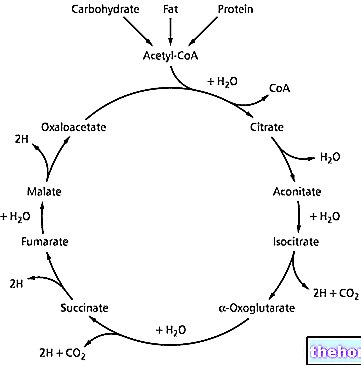
Encim, ki katalizira četrto stopnjo krebsovega cikla, je α-keto glutarat dehidrogenaza; ta encim je encimski kompleks, zelo podoben piruvat dehidrogenazi. Oba sta sestavljena iz 48-60 beljakovin, v katerih so prepoznane tri različne encimske aktivnosti in imajo tudi enake encimske kofaktorje; so zelo podobni encimi, ker delujejo na podobnih substratih: tako piruvat kot l "α-keto glutarat, so α- keto kisline. Mehanizem delovanja obeh encimskih kompleksov je enak.
Napad tiamin pirofosfata na karbonil (C = O) "α-ketoglutarat, vodi do njegove dekarboksilacije in nastane karboksihidroksi propil derivat. Z naknadnim prenosom v lipoamid poteka notranji redoks proces, iz katerega dobimo derivat lipoamid karboksi ali sukcinil lipoamid.
Sukcinil lipoamid nato reagira s koencimom A, da nastane sukcinil koencim A (ki se nadaljuje v Krebsovem ciklu) in reducirani lipoamid, ki se s FAD ponovno oksidira: nastali FADH2 se ponovno oksidira z NAD + in NADH. Na tej stopnji je torej prišlo do druge odstranitve ogljika iz ogljikovega ogrodja v obliki ogljikovega dioksida.
Acilna skupina, povezana s koencimom A, je v aktivirani obliki, to je z visoko vsebnostjo energije: zato je mogoče izkoristiti energijo sukcinilnega koencima A.

V peti fazi Krebsovega cikla je sukcinil koencim A izpostavljen delovanju sukcinil tiokinaza; o njegovem načinu delovanja sta bili postavljeni dve hipotezi: opisali bomo le eno od obeh, ker je najbolj akreditirana. V skladu s to hipotezo sukcinil koencim A napada dušik histidina (Hys) encima: sprošča se koencim A in nastane adukt, ki izhaja iz histidina, kot vmesni produkt, to je sukcinil-encim (ali sukcinil-Hys ); ortofosfat deluje na ta vmesni produkt, kar vodi v sproščanje sukcinata in nastanek fosfoencima. Z vidika energije GTP = ATP: vez, ki zagotavlja energijo, je pri obeh vrstah enaka (to je anhidridna vez med fosforil Β in fosforil γ). V nekaterih primerih se GTP uporablja kot material z visoko vsebnostjo energije, običajno pa se GTP pretvori v ATP z delovanjem encima nukleozid difosfo kinaza (NDPK); je encim, ki ga najdemo v celicah in katalizira naslednjo reakcijo:
N1TP + N2DP → N1DP + N2TP
Generični NiTP ® nukleozid trifosfat
Generični NiDP ® nukleozid difosfat
To je reverzibilna reakcija; v našem primeru se zgodi:
GTP + ADP → BDP + ATP
zato lahko nadaljuje v desno ali levo tudi pri majhnih nihanjih koncentracij reagentov.
Če Krebsov cikel teče s takšno hitrostjo, da povzroči proizvodnjo ATP, ki je višja od energetske potrebe, je ADP malo, medtem ko je veliko ATP: reakcija, ki jo katalizira nukleozid difosfokinaza, je usmerjeno v levo (GTP se kopiči, če nukleozid difosfo kinaza nima dovolj substrata, to je ADP). GTP je torej signal razpoložljivosti energije in zato upočasnjuje krebsov cikel.
Šesta stopnja krebsovega cikla vodi do tvorbe fumarata z delovanjem sukcinat dehidrogenaza; ta encim daje stereospecifično reakcijo, saj vedno nastane nenasičen (to je alken) trans, to je fumarat (medtem ko je cis izomer maleat). Sukcinat dehidrogenaza se nahaja na notranji mitohondrijski membrani, medtem ko so vsi drugi encimi v krebsovem ciklu razpršeni po celotni mitohondriji.
Sukcinat dehidrogenaza ima kot kofaktor FAD; zavira ga oksaloacetat (inhibicija povratne zveze), medtem ko ima sukcinat in fumarat kot pozitivni modulator (aktivator). Krebsov cikel potrebuje energijo, zato je edina možnost, da dobimo oksaloacetat pri pacientu, ta, da je njegova koncentracija zelo visoka: malat je eden od presnovkov z najvišjo koncentracijo v celicah. dejstvo, da koncentracija oksaloacetata zaradi delovanja citrat sintaze ohranja nizko. Reakcija, ki jo katalizira sukcinat dehidrogenaza, je torej samohranilna reakcija in to je edini način za pretvorbo malata v oksaloacetat.
Koncentracija mitohondrijskega malata mora biti združljiva s koncentracijo citoplazmatskega malata: samo če je koncentracija mitohondrijskega malata tako visoka, da zagotavlja pretvorbo malata v oksaloacetat (v Krebsovem ciklu), se lahko malat uporabi tudi v drugi načini (ki so citoplazmatski): v citoplazmi se lahko malat pretvori v oksaloacetat, iz katerega lahko dobimo aspartat z delovanjem GOT (je transaminaza) ali glukoze z glukoneogenezo.
Vrnemo se na sedmo stopnjo krebsovega ciklusa, ki ga katalizira encim fumarasi: voda nastane na stereospecifičen način, da nastane L-malat.
V zadnji fazi Krebsovega cikla, o katerem smo že govorili, se dogaja malat dehidrogenaza. Ta encim uporablja molekulo NAD + za svoje katalitično delovanje.
Tako smo zaključili opis različnih stopenj krebsovega cikla.
Krebsov cikel je popolnoma reverzibilen.
Za povečanje hitrosti krebsovega cikla je mogoče povečati koncentracijo presnovkov, prisotnih v tem ciklu; ena od strategij za povečanje hitrosti krebsovega cikla je pretvorba dela piruvata, ki vstopa v mitohondrije, v oksaloacetat (z delovanjem piruvat karboksilaze) in ne pretvarjanje vsega v acetil koencim A: s tem se poveča koncentracija oksaloacetata, ki je presnovek Krebsovega cikla in zato povečuje hitrost celotnega cikla.
V krebsovem ciklu se tri NAD + pretvorijo v tri NADH in en FAD v FADH2, poleg tega pa se pridobi GTP: s preusmerjanjem redukcijske moči, pridobljene s krebsovim ciklom, se proizvede nadaljnji ATP; v dihalni verigi se redukcijska moč prenaša iz NADH in FADH2 v kisik: ta prenos je posledica vrste encimov, ki se nahajajo na mitohondrijski membrani in ki v svojem delovanju vodijo v proizvodnjo ATP.
Procesi dihalne verige so eksergonski procesi in sproščena energija se uporablja za proizvodnjo ATP; namen celice je izkoristiti eksergonske procese za sintezo ATP. Za vsako molekulo NADH, ki vstopi v dihalno verigo, dobimo 2,5 molekule ATP in za vsako FADH2 1,5 molekule ATP; ta raznolikost je posledica dejstva, da FADH2 vstopi v dihalno verigo na nižji ravni kot NADH.
Z zmanjšanjem moči aerobne presnove dobimo 30-32 ATP (219-233 kcal / mol) z izkoristkom približno 33% (učinkovitost anaerobne presnove je približno 2%).